Groupe 4 : Quel est l’effet de l’éclairage nocturne par des lampadaires sur les végétaux en ville?
Vesna Ignjatovic : vesna.ignjatovic@etu.sorbonne-universite.fr
Caroline Jung : caroline.jung@etu.sorbonne-universite.fr
Zelda Lefeuvre : zelda.lefeuvre@etu.sorbonne-universite.fr
Introduction :
Date de début : 22/10/2024
Date de fin estimée : 29/11/2024
À Paris, on compte plus de 171 000 lampadaires qui éclairent toute la nuit la ville (1). S’ils permettent aux habitants de la « ville-lumière » de se repérer, même la nuit, ces éclairages artificiels perturbent le reste des êtres vivants. Ces lampadaires constituent de la pollution lumineuse, définie comme la lumière artificielle qui perturbe l’alternance du jour et de la nuit sur le cycle de 24h (rythme nycthéméral) dans les écosystèmes. Les activités anthropiques occupant de plus en plus de territoire, la pollution lumineuse augmente de sorte qu’aujourd’hui, 88 % de l'Europe est éclairé la nuit (2). Depuis les années 90, ce phénomène commence à inquiéter et à ce jour, 25% de la population mondiale ne peut plus apercevoir la voie lactée (3). De nombreuses études ont déjà montré l’effet de la lumière nocturne sur des animaux, notamment sur l’orientation des oiseaux migrateurs, les relations proie-prédateur, etc. Les conséquences sur les végétaux sont moins étudiées, pourtant, elles n’en restent pas moins notables.
Notamment, les végétaux sont dépendants pour leur croissance et leur développement de la lumière et ils ne font pas la différence entre la lumière naturelle et la lumière artificielle. L’éclairage nocturne déstabilise ainsi le cycle naturel de lumière en augmentant la durée du jour pour la plante. Il affecte ainsi la germination, la croissance, l’expansion des feuilles, la floraison, le développement des fruits et la sénescence (4). En effet, plus qu’une ressource, la lumière est aussi un indicateur biologique pour les plantes. C'est l’obscurité lors du cycle de 24 heures qui assure les processus de développement tels que la dormance, la croissance des pousses et la floraison (3). Les plantes à jour court par exemple, fleurissent à l’automne quand la durée du jour diminue. En effet, elles profitent des longues nuits pour entamer leur processus de floraison. C’est aussi l’augmentation de la durée de la nuit, qui permet l'initiation du processus de « dormance » assurant à la plante de résister au froid de l'hiver. Or la lumière artificielle envoie des fausses indications aux plantes photosensibles.
Il est donc important d’évaluer les effets de l’éclairage nocturne sur les végétaux, d’autant plus que de nombreux animaux en sont dépendants. Une perturbation du cycle de la plante pourrait avoir des effets en cascade se répercutant sur les herbivores, puis les insectivores, etc (2). Par exemple, si les fleurs fleurissent trop tôt, elles seront déjà fanées lorsque les insectes se mettront en quête de leur nectar (5).
Nous allons ainsi nous demander quel est l’effet de l’éclairage nocturne par des lampadaires sur les végétaux en ville. Nous étudierons précisément les conséquences d’une exposition prolongée à la lumière artificielle sur les radis. Deux hypothèses se présentent : soit une exposition prolongée durant la nuit n’a pas d’effet sur le développement des plantes (H0) soit une exposition prolongée durant la nuit a un effet sur le développement des plantes (H1). Nous nous attendons à ce que l’éclairage par les lampadaires ait un effet sur la croissance des plantes. En ce sens, Guo et al ont montré que le radis grossissait à mesure que la durée d’éclairement augmentait (6). L’intensité lumineuse semble ne pas être suffisante pour impacter la photosynthèse, en revanche, elle devrait perturber le rythme biologique de la plante.
I. Matériel et méthodes
Nous avons choisi comme modèle le radis, Raphanus sativus, plus précisément la variété hybride F1 qui pousse très rapidement. Le radis est une plante modèle appréciée, utilisée pour de nombreuses études comme sur les réponses aux polluants atmosphériques, à la température, à l’apport de lumière ou encore au stress hydrique. Cette plante s’avère être parfaite pour notre projet en raison de sa petite taille, facilement quantifiable, qui permet d’avoir un nombre important de réplicats et de facilement analyser le phénotype et l’allocation des ressources (7). De plus, son cycle de vie est rapide (une vingtaine de jours), ce qui rentre dans le temps qui nous est imparti pour notre projet. Pour ce qui est de la photosensibilité du radis, plusieurs études nous confirment que ce modèle est bien affecté par la durée d’exposition à la lumière. Enfin, les radis que nous avons choisi sont des hybrides F1, ce qui nous permet d’avoir moins de variations entre les individus et que chaque groupe soit donc affecté de la même manière : ils sont synchronisés et leurs phénotypes sont assez homogènes.
Pour évaluer l’impact de la durée d’éclairement, nous répartissons les plantes en 3 groupes dans lesquels la durée d’exposition est plus ou moins longue. Notre premier groupe, contrôle, se constitue des plantes exposées la journée (11h). Le deuxième, correspondant aux conditions de Paris, regroupe des plantes exposées jour et nuit (24h). Enfin, les plantes du troisième groupe sont exposées la journée et la moitié de la nuit (17h). Des programmateurs sont utilisés.
Dans chacun des groupes, il y a 10 plantes soit 30 plants au total. Nous isolons les groupes avec des panneaux en cartons placés entre eux pour limiter les échanges de lumière. Nous utilisons des lampes LED blanches neutres pour imiter les lampadaires parisiens. En effet, la majorité des lampadaires utilisés dans les villes sont des luminaires LED produisant une lumière à “spectre complet” (1) soit compris entre 300-700 nanomètres, plutôt blanc neutre ou chaud. Nous avons choisi d’utiliser une lampe LED blanc neutre, soit une température de 4000 K, éclairage se rapprochant le plus de la température de la lumière des étoiles jaunes, comme le soleil (8). Comme la plupart des plantes, dont les radis, utilisent majoritairement les longueurs d’ondes dans le bleu (380-480 nm) et dans le rouge (620-700 nm), un spectre complet de lumière blanche sera efficace pour faire pousser nos radis (9). De plus, les lampes LED ont l’avantage de ne pas trop s’échauffer.
Pour limiter les biais, nous avons tiré au hasard les graines, contrôlé la quantité d’eau donnée à chaque plante et la profondeur à laquelle les graines sont plantées. Afin de s’assurer que nos plantes germent, nous avons planté deux graines dans chaque pot. Si les deux germent, nous en retirons une pour la suite de l’expérience afin d’éviter le phénomène de compétition.
Pour finir, concernant notre protocole, nous plantons les graines de radis en semaine 1. Tous les deux jours, nous arrosons nos plantes au besoin (10) et suivons leur phénotype. Nous mesurons au cours du temps plusieurs variables dépendantes quantitatives y, comme la vitesse de tubérisation, le nombre de feuilles, la surface des feuilles grâce au logiciel EasyLeafArea. A la fin (31 jours après la mise en pot), nous pèserons les masses fraîches et sèches aériennes et souterraines des plantes, et le diamètre des radis. La variable indépendante x que nous ferons varier est la durée de l’exposition à la lumière (quantitatif). Nous analyserons ces divers données à l’aide de Rstudio, vous trouverez les scripts en annexe.
Matériaux / Outils / Machines :
-
-
30 pots en plastique de 7 cm de diamètre
-
3 plateaux pour poser les pots
-
3 panneaux LED blanc 4000 K
-
2 programmateurs pour les lumières
-
2 planches de cartons pour séparer les 3 lots de plantes
-
60 graines de radis
-
Du terreau
-
Une barre en métal pour suspendre les lampes à la même hauteur
-
Construction :

Journal de bord :
22/10 : nous avons planté les radis, 2 graines dans chaque pot, afin d’assurer un maximum de germination et nous les avons arrosées avec 700 mL par plateau.
24/10 : arrosage des plantes avec 500 mL par plateau.
28/10 : arrosage des groupes (300 mL par groupe), calcul de la surface foliaire
30/10 : arrosage des groupes (200 mL par groupe), calcul de la surface foliaire puis déracinage d’une plante dans les pots où il y en avait 2 pour éviter la compétition
4/11 : arrosage des groupes (300 mL par groupe), calcul de la surface foliaire
6/11 : arrosage des groupes (500 mL par groupe), calcul de la surface foliaire. Ci-après, des photos de nos groupes dans 3 conditions après 2 semaines de culture (de gauche à droite : groupe de 17h, groupe de 24h, groupe de 11h) :



8/11 : arrosage des groupes (400 mL par groupe), calcul de la surface foliaire
12/11: arrosage de 700ml et calcul de la surface foliaire
15/11: arrosage de 300 mL et calcule de la surface foliaire
18/11: arrosage de 700 mL et calcule de la surface foliaire
II. Résultats :
Vitesse de tubérisation : Nous avons observé la présence d’un radis dès le 13ème jour chez le groupe de 24h, au 17ème jour chez le groupe de 17h et au jour 21 chez le groupe de 11h.
Observations qualitatives (Figure 1) : Le groupe de 24h avait toujours besoin d’eau plus vite que les deux autres groupes (la terre était toujours sèche quand on venait les arroser). Nous avons observé quelques chloroses et nécroses chez le groupe 24h mais pas chez les deux autres. Souvent, ce même groupe avait des feuilles flétries.
Nombre de feuilles (Figure 2) : Le 31ème jour, les plantes exposées 24h par jour à la lumière ont significativement moins de feuilles que les plantes contrôles, exposées 11h à la lumière ( p-value: 3.688e-08). Il y a une baisse du nombre de feuilles chez le groupe de 24h autour de 17ème jour, non observé chez les autres groupes.
Surface des feuilles (Figures 3,4) : La surface évolue de manière similaire jusqu’au jour 17 auquel on note une divergence entre le groupe de 24h et ceux de 11h et 17h. La surface du groupe de 24h diminue, cela va dans le sens des résultats précédents avec la perte de feuilles de ce groupe. Au 31ème jour, la surface des feuilles des plantes exposées 24h par jour à la lumière est significativement inférieure à celle des plantes exposées 11h par jour (p-value = 2.736e-10). Il n’y a pas de différence significative entre les groupes de 11h et 17h.
Masse fraîche aérienne (Figure 5) : Les plantes exposées 24h par jour à la lumière ont une masse fraîche aérienne significativement inférieure à celle des plantes exposées 11h par jour (p-value = 3.77e-09). Il n’y a pas de différence significative entre les groupes de 11h et 17h.
Masse fraîche souterraine (Figure 6) : Il n’y a pas de différence significative entre les différents groupes (p-value = 0.57).
Ratio masse fraîche A/S (Figure 7) : Les plantes exposées 24h et 17h par jour à la lumière ont un ratio de masse fraîche A/S significativement inférieure à celui des plantes exposées 11h par jour (p-value: 0.0002383).
Masse sèche aérienne (Figure 8) : Il n’y a pas de différence significative de la masse sèche aérienne entre les différents groupes (p-value = 0.697).
Masse sèche souterraine (Figure 9) : La masse sèche des radis des plantes exposées 24h par jour à la lumière est significativement plus élevée que celle des radis des plantes exposées 11h par jour (p-value = 1.013e-05). Il n’y a pas de différence significative entre les plantes de 11h et 17h.
Ratio masse sèche A/S (Figure 10) : Les plantes exposées 24h par jour au soleil ont un ratio masse sèche A/S significativement inférieur à celui des plantes exposées 11h par jour (p-value = 0.0242). Il n’y a pas de différence significative entre les groupes de 11h et 17h.
Ratio masse fraîche/masse sèche (Figure 11) : Les plantes exposées 24h par jour à la lumière ont un ratio MF/MS significativement inférieur à celui des plantes exposées 11h par jour (p-value = 0.00024). Il n’y a pas de différence significative entre les plantes de 11h et 17h.
Diamètre radis (Figure 12) : Il n’y a pas de différences significatives entre les diamètres des radis des différents groupes (p-value = 0.11).
Surface spécifique (Figure 13) : La surface spécifique des plantes du groupe de 24h est significativement inférieure à celle des plantes du groupe de 11h (p-value = 3.688e-08). Il n’y a pas de différence significative entre les plantes de 11h et 17h.
III. Discussion
L’accroissement de l’utilisation de lampadaires la nuit nous a amené à étudier l’effet de l’éclairage artificiel sur les végétaux. Nos résultats confirment la perturbation des végétaux par une durée prolongée d’exposition à la lumière. L’effet se note à plusieurs niveaux : des effets délétères sur la plante tels que de la nécrose, du stress hydrique ; des perturbations du cycle biologique comme une tubérisation précoce ; des effets sur la croissance aérienne et souterraine.
Nous voyons que le rapport masse fraîche aérienne/masse fraîche souterraine (MFa/MFr) est supérieur chez les plantes les moins éclairées (11h) par rapport aux deux autres groupes. Nous pouvons supposer que la réduction du rapport est un effet de l’éclairement prolongé : en effet, nous avons vu que les plantes du groupe de 24h ont une surface foliaire, un nombre de feuilles et une masse fraîche aérienne significativement moins importants que les deux autres groupes, ce qui montre qu’elles ont tendance à moins développer ou à réduire leur surface aérienne, trop exposée à la lumière, puisqu’elles en ont moins besoin. En parallèle, elles vont donc allouer plus de ressources à l’appareil racinaire afin de maximiser l'absorption de nutriments et notamment d’eau, qui comme nous le verrons ci-dessous, peut se faire plus rare dans des conditions d’éclairement prolongé.
Ainsi, la masse fraîche racinaire augmente tandis que la masse fraîche aérienne diminue, ce qui provoque la diminution du rapport MFa/MFr. Nous observons par ailleurs les mêmes résultats pour le rapport des masses sèches (MSa/MSr).
En comparant les masses aériennes fraîches et sèches, nous pouvons voir qu’il y a une différence significative seulement pour la masse fraîche : le groupe de 24h utilise plus d’eau que les deux autres, provoquant chez lui un stress hydrique, il y a donc moins d’eau dans ses feuilles, et c’est pour cela qu’une fois séchées, nous n'observons plus de différence.
Du côté de la masse sèche souterraine (MSr), nous observons bien des valeurs plus élevées chez les radis éclairés 24h par rapport aux autres groupes, qui étaient moins soumis au stress hydrique. Pour ce qui est de l’absence de différence entre les 3 groupes pour la masse fraîche, nous supposons que comme pour les feuilles, les radis des groupes de 11h et 17h étaient simplement plus gorgés d’eau pour cette raison.
Ainsi, nous concluons qu’une exposition fortement prolongée à la lumière permet de favoriser la croissance souterraine des radis.
En revanche, nous ne notons pas de différences significatives pour le diamètre des radis de chacun des groupes. C’est pourtant un résultat auquel on aurait pu s’attendre, qui avait d’ailleurs été trouvé par Combe, L., et al. lors de leur étude en 1988 (11). Ces derniers avaient également montré que la tubérisation et la germination sont déclenchées après un certain nombre d’heures d’éclairement. Ainsi, des radis éclairés plus longtemps dans la journée devraient atteindre ce seuil avant les autres. En ce sens, nous avons bien observé que les radis sont apparus en premier chez le groupe de 24h après 13 jours d’éclairement non stop soit 312 heures d’éclairement, suivi du groupe de 17 heures après 289 heures accumulées d’exposition à la lumière (au bout de 17 jours). Il semblerait donc bien que la tubérisation soit précoce chez les plantes exposées continuellement à une lumière artificielle. Ce résultat démontre conséquemment qu’un éclairage nocturne continu peut perturber les cycles biologiques naturels des végétaux.
Par ailleurs, les plantes exposées constamment à la lumière présentent des signes de stress. Nous avons pu observer des lésions et des chutes de feuilles uniquement chez le groupe de 24h. Bennie, Jonathan, et al. ont écrit en 2016 qu’une exposition constante à la lumière conduit à une augmentation des lésions foliaires dues à l’ozone (12). On peut émettre l’hypothèse que face à un éclairement trop important, les plantes de 24h ont produit des espèces réactives à oxygène menant à des lésions. Mais des expériences complémentaires devraient être réalisées pour le prouver. De plus, nous avons observé que les plantes de 24h ont un ratio masse fraîche sur masse sèche (MF/MS) significativement inférieur à celui des plantes de 11h. La mesure de ce ratio permet d’indiquer le contenu en eau d’une plante. Les plantes éclairées constamment ont un ratio plus faible ce qui signifie qu’elles ont une plus faible quantité d’eau par rapport à leur matière sèche. Ceci est observé chez les plantes en stress hydrique. Les plantes ayant un faible ratio sont souvent mieux adaptées à la sécheresse voire résistantes (13). A l’inverse un ratio élevé est plutôt signe d’une croissance rapide et une forte biomasse végétale. Les plantes de 11h et 17h ont un effet une masse fraîche supérieure à celle des plantes de 24h. Nous avons par ailleurs mesuré la surface spécifique foliaire (SSF) et montré que les plantes exposées 24h par jour au soleil ont une surface spécifique inférieure aux deux autres groupes. NIRY Fenosoa Sylvia explique dans son étude sur la SSF menée en 2010 qu’une SSF élevée est liée à une croissance rapide de la plante, qui est dépendante de l’activité photosynthétique et de la respiration de la biomasse aérienne (14). Ainsi, nous pouvons déduire que les plantes de 24h ont une activité photosynthétique plus faible car elles ont une biomasse aérienne plus faible et qu’elles investissent moins de matière dans la photosynthèse. On suppose également que la SSF est plus faible car leurs feuilles sont plus épaisses et étroites à cause de l’éclairage constant et d’un potentiel stress hydrique.
Enfin, on peut noter que pour la plupart de nos résultats, les différences étaient significatives entre les groupes de 11h et 24h mais non entre les groupes de 11h et 17h. On peut ainsi supposer qu'éteindre les lampadaires la moitié de la nuit à Paris réduirait en grande partie leurs effets néfastes sur les végétaux. Toutefois, nos résultats ne permettent pas entièrement de conclure quant à l’utilisation de lampadaires la nuit car les végétaux n’étaient pas disposés à la même distance de la lumière artificielle qu'en ville et donc éclairés par la même intensité lumineuse. Bien que probablement accentués, ces résultats nous donnent tout de même une idée des éventuels dommages d’un éclairage nocturne constant.
Pour aller plus loin, nous aurions pu mettre en place un autre protocole jouant cette fois non pas sur la photopériode mais plutôt sur l’intensité de l’éclairement nocturne. Nous aurions également pu mesurer la teneur en chlorophylle des feuilles, ainsi qu’en espèces réactives à l’oxygène.
BIBLIOGRAPHIE
-
Team, Eclairage pour Tous :. Tendances et Guides pour l’Éclairage Intérieur et Extérieur. « Eclairage Public à Paris : Défis, Solutions et Innovations ». Eclairage pour Tous : Tendances et Guides pour l’Éclairage Intérieur et Extérieur, https://www.eclairagepublic.org/2024/02/eclairage-public-paris.html. Consulté le 20 octobre 2024.
-
Gracci, Fiorenza. « Quel est l’impact de l’éclairage public sur la vie des arbres ? » Science et vie, 31 octobre 2019, https://www.science-et-vie.com/questions-reponses/quel-est-limpact-de-leclairage-public-sur-la-vie-des-arbres-8714.html. Consulté le 20 octobre 2024.
-
William R. Chaney. 2002. Does Night Lighting Harm Trees? Department of Forestry and Natural Resources.https://www.extension.purdue.edu/extmedia/fnr/fnr-faq-17.pdf
-
BRIGGS W.R. (2006).- Physiology of Plant Responses to Artificial lighting Pp. 389-411 in Ecological consequences of artificial night lighting (C. RICH and T. LONGCORE, eds.).
-
PETITJEAN, IRIS. « Les arbres aussi veulent dormir ». le journal minimal, 3 avril 2018, https://lejournalminimal.fr/arbres-veulent-dormir/. Consulté le 20 octobre 2024.
-
Guo, Rongfang, et al. « Effect of photoperiod on the formation of cherry radish root ». Scientia Horticulturae, vol. 244, janvier 2019, p. 193‑99. ScienceDirect, https://doi.org/10.1016/j.scienta.2018.09.044.
-
Kostka-Rick, Reinhard, et William J. Manning. « Radish (Raphanus sativus L.): A model for studying plant responses to air pollutants and other environmental stresses ». Environmental Pollution, vol. 82, no 2, janvier 1993, p. 107‑38. ScienceDirect, https://doi.org/10.1016/0269-7491(93)90109-2.
-
Avantages de l’éclairage public à LED : https://www.aecilluminazione.fr/avantages-de-leclairage-public-exterieur-a-led/. Consulté le 23 octobre 2024.
-
The Ideal LED Grow Light Spectrum for Plants : https://bioslighting.com/horticulture-blog/grow-light-spectrum-led-plants/. Consulté le 23 octobre 2024.
-
The Effect of Light Intensity on the Height of Radish Plants (Raphanus sativus) https://www.tsfx.com.au/wp-content/uploads/2018/03/Year-12-Biology-A.pdf. Consulté le 23 octobre 2024.
-
Combe, L., et al. « Croissance et tubérisation du radis ( Raphanus sativus ) selon la durée d’éclairement ». Canadian Journal of Botany, vol. 66, no 9, septembre 1988, p. 1834‑40. DOI.org (Crossref), https://doi.org/10.1139/b88-250.
-
Bennie, Jonathan, et al. « Ecological Effects of Artificial Light at Night on Wild Plants ». Journal of Ecology, édité par Nathan Swenson, vol. 104, no 3, mai 2016, p. 611‑20. DOI.org (Crossref), https://doi.org/10.1111/1365-2745.12551.
-
Pérez-Harguindeguy, N., et al. « Corrigendum to: New Handbook for Standardised Measurement of Plant Functional Traits Worldwide ». Australian Journal of Botany, vol. 64, no 8, 2016, p. 715. DOI.org (Crossref), https://doi.org/10.1071/BT12225_CO.
-
NIRY, et al. 2010. Détermination de la surface spécifique foliaire (SSF), relation avec le fonctionnement physiologique chez dix variétés de riz pluvial. https://open-library.cirad.fr/files/4/1648__Feno_2010.pdf
ANNEXES
Scripts R :
radis <- read_excel("radis.xlsx", range = "A1:Q301", na = "NA")
radis$Durée_lumière <- as.factor(radis$Durée_lumière)
View(radis)
Nbfeuillesmodele <- glm(Nbfeuilles ~ Durée_lumière, family = "poisson", data = radis)
summary(mon_modele)
anova(Nbfeuillesmodele,test="Chisq")
plot(Nbfeuilles ~ jitter(Jour), data = radis,
col = as.factor(radis$Durée_lumière),
xlab = "jours", ylab = "Nombre de feuilles")
mon_modele <- lm(Surface ~ Durée_lumière, data = radis)
summary(mon_modele)
anova(mon_modele,test="Chisq")
plot(Surface ~ jitter(Jour), data = moy_radis,
col = as.factor(moy_radis$Durée_lumière),
xlab = "jours", ylab = "Surface")
radis31 <- read_excel("radis31.xlsx", range = "A1:Q31")
radis31$Durée_lumière <- as.factor(radis$Durée_lumière)
View(radis31)
mon_modele <- lm(MFAérienne ~ Durée_lumière, data = radis)
summary(mon_modele)
anova(mon_modele,test="Chisq")
boxplot(MFAérienne ~ Durée_lumière, data = radis,
xlab = "jours", ylab = "Masse fraîche aérienne")
mon_modele <- lm(RatioMFA_R ~ Durée_lumière, data = radis31)
summary(mon_modele)
anova(mon_modele,test="Chisq")
boxplot(RatioMFA_R ~ Durée_lumière, data = radis,
xlab = "jours", ylab = "Ratio masse fraîche aérienne sur masse fraîche du radis")
mon_modele <- lm(MSradis ~ Durée_lumière, data = radis31)
summary(mon_modele)
anova(mon_modele,test="Chisq")
boxplot(MSradis ~ Durée_lumière, data = radis31, xlab = "jours", ylab = "Masse sèche du radis")
mon_modele <- lm(MSAérienne ~ Durée_lumière, data = radis31)
summary(mon_modele)
anova(mon_modele,test="Chisq")
boxplot(MSAérienne ~ Durée_lumière, data = radis31, xlab = "jours", ylab = "Masse sèche aérienne")
mon_modele <- lm(RatioMSA_R ~ Durée_lumière, data = radis31)
summary(mon_modele)
boxplot(RatioMSA_R ~ Durée_lumière, data = radis31, xlab = "jours", ylab = "Ratio masse aérienne sur masse radis")
mon_modele <- lm(RatioMF_MS ~ Durée_lumière, data = radis31)
summary(mon_modele)
boxplot(RatioMF_MS ~ Durée_lumière, data = radis31, xlab = "jours", ylab = "Ratio masse fraîche sur masse sèche")
mon_modele <- lm(Surface ~ Durée_lumière, data = radis31)
summary(mon_modele)
anova(mon_modele,test="Chisq")
mon_modele <- lm(Diamètre_radis ~ Durée_lumière, data = radis31)
summary(mon_modele)
anova(mon_modele,test="Chisq")
boxplot(Diamètre_radis ~ Durée_lumière, data = radis31, xlab = "jours", ylab = "Diamètre du radis")
mon_modele <- lm(Surface_spécifique ~ Durée_lumière, data = radis31)
summary(mon_modele)
anova(mon_modele,test="Chisq")
boxplot(Surface_spécifique ~ Durée_lumière, data = radis31, xlab = "jours", ylab = "Surface spécifique")
Résultats :

Figure 1 : Plante du groupe de 24h présentant des nécroses et chloroses (18/11, jour 27)
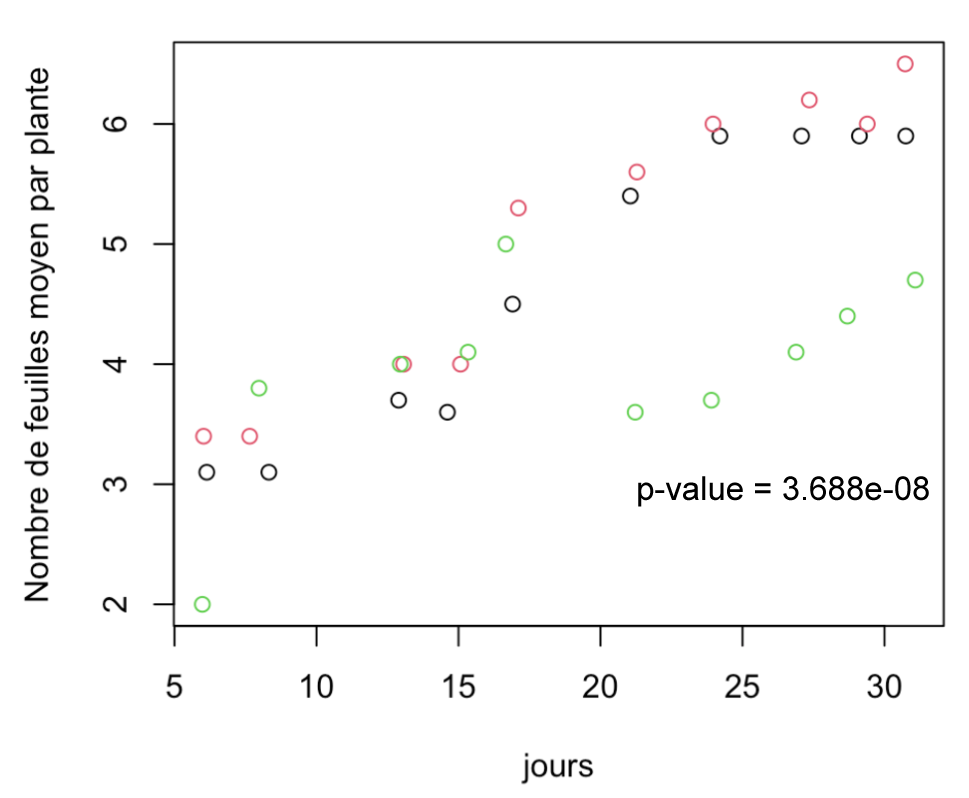
Figure 2 : Evolution du nombre de feuilles des plantes de chaque groupe (11h, 17h et 24h) au cours du temps
rouge = 11h, noir = 17h, vert = 24h
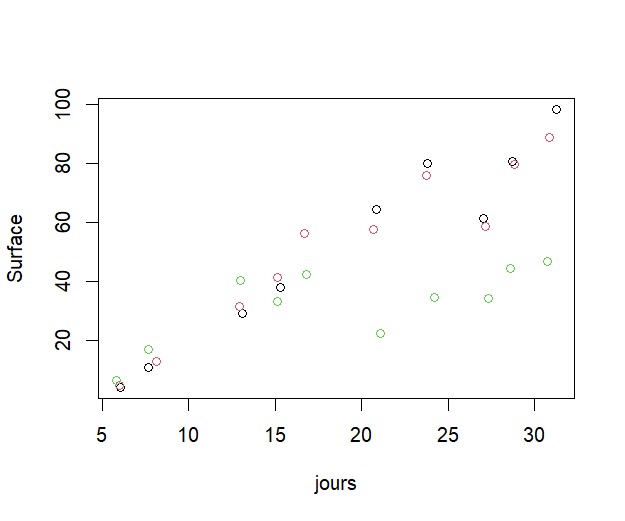
Figure 3 : Evolution de la surface foliaire des plantes de chaque groupe (11h, 17h, 24h) au cours du temps
rouge = 11h, noir = 17h, vert = 24h
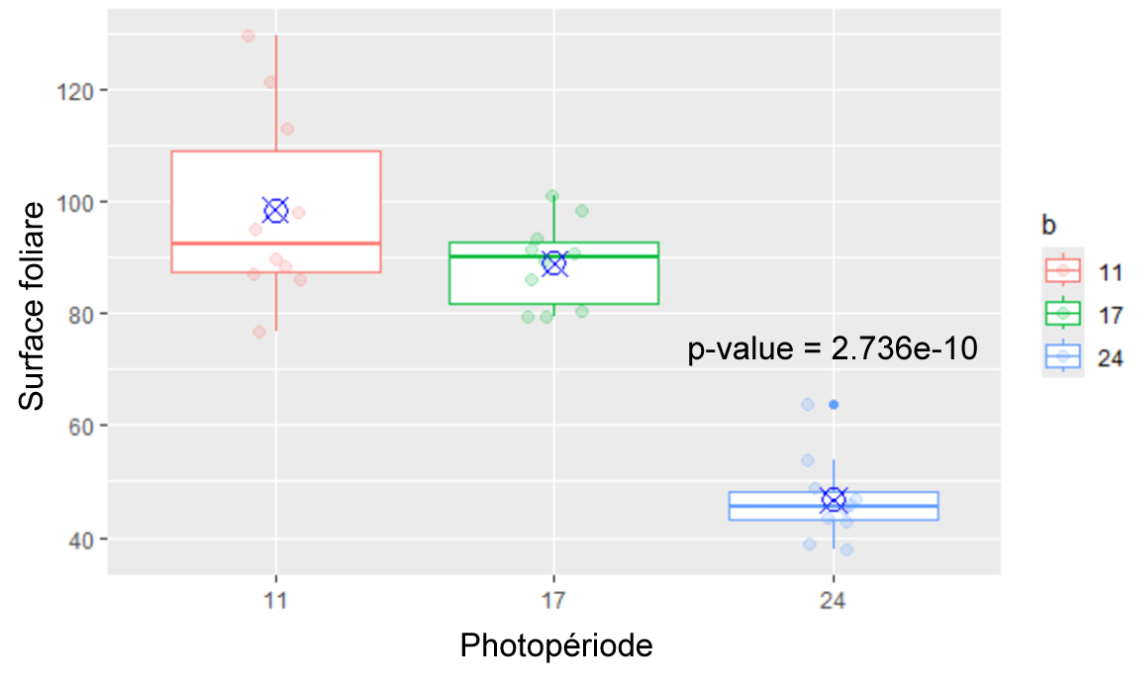
Figure 4 : Surface foliaire des plantes des différents groupes (11h, 17h, 24h) au 31ème jour
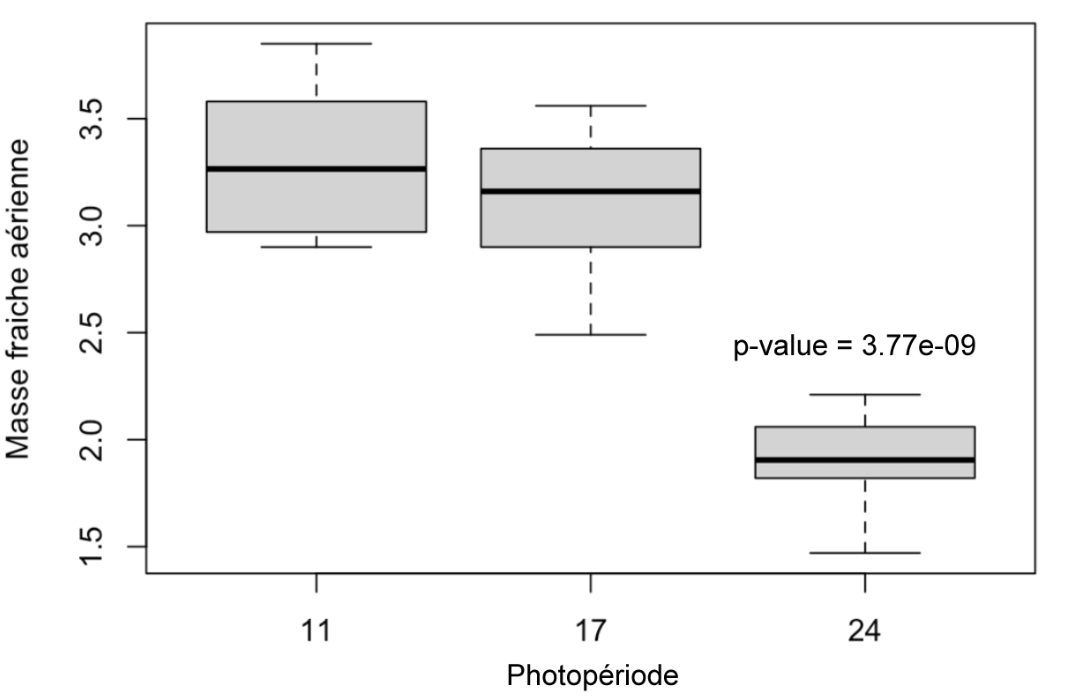
Figure 5 : Masse fraîche aérienne des plantes des différents groupes (11h, 17h, 24h) mesurées au 31ème jour
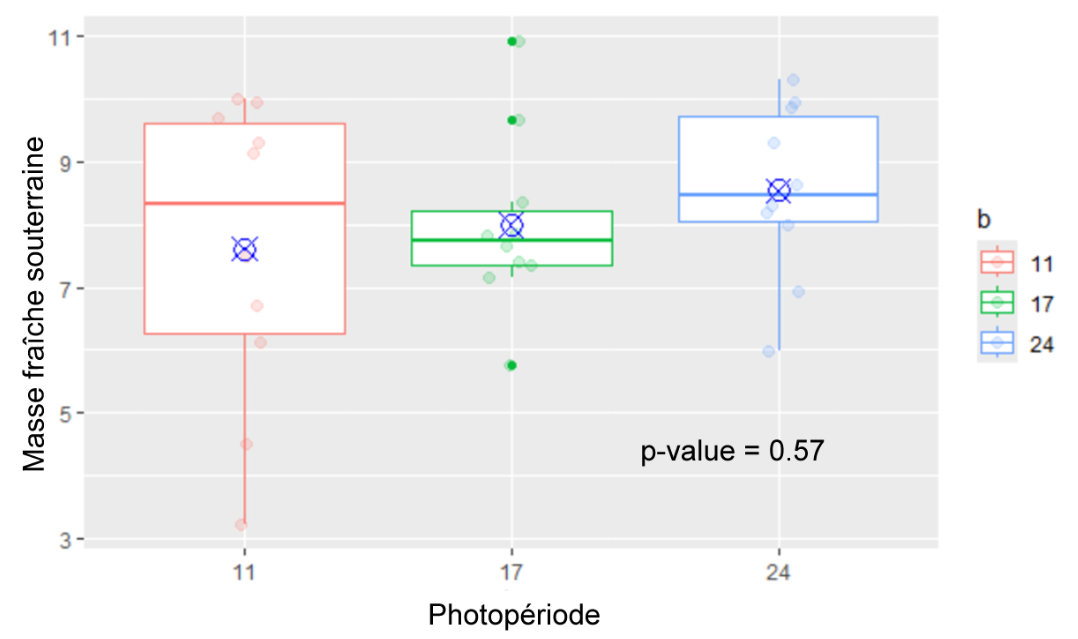
Figure 6 : Masse fraîche souterraine des plantes des différents groupes (11h, 17h, 24h) mesurées au 31ème jour
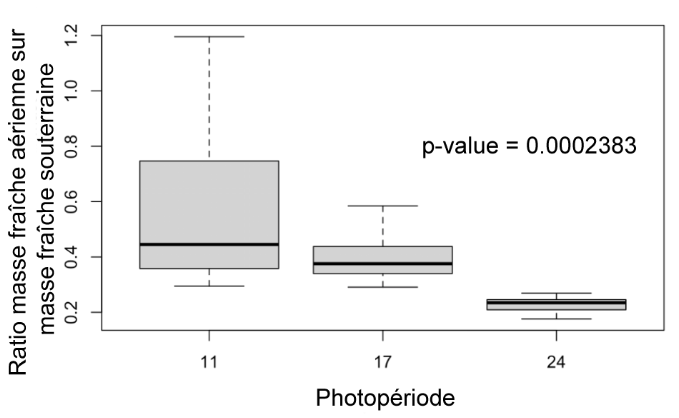
Figure 7 : Ratio des masses fraîches aériennes sur souterraines des plantes des différents groupes (11h, 17h, 24h) au 31ème jour
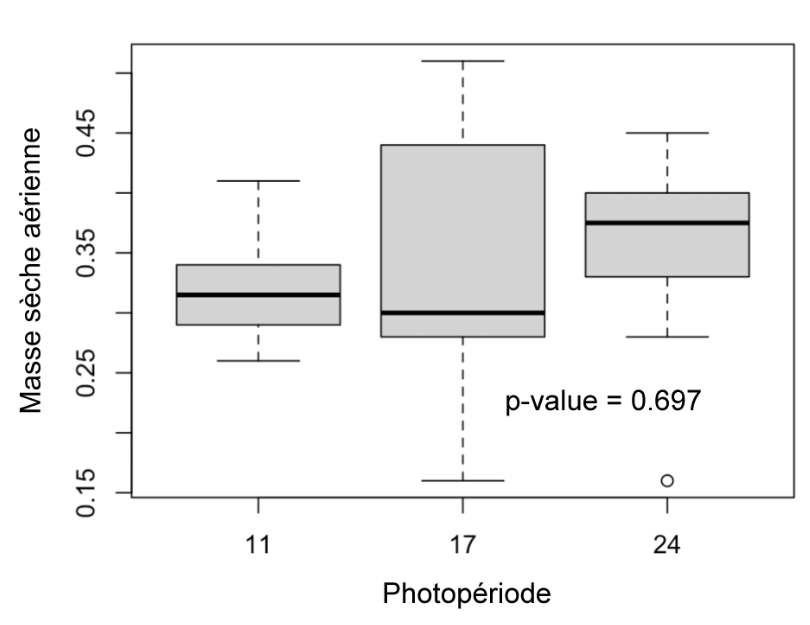
Figure 8 : Masse sèche aérienne des plantes des différents groupes (11h, 17h, 24h) mesurées au 31ème jour
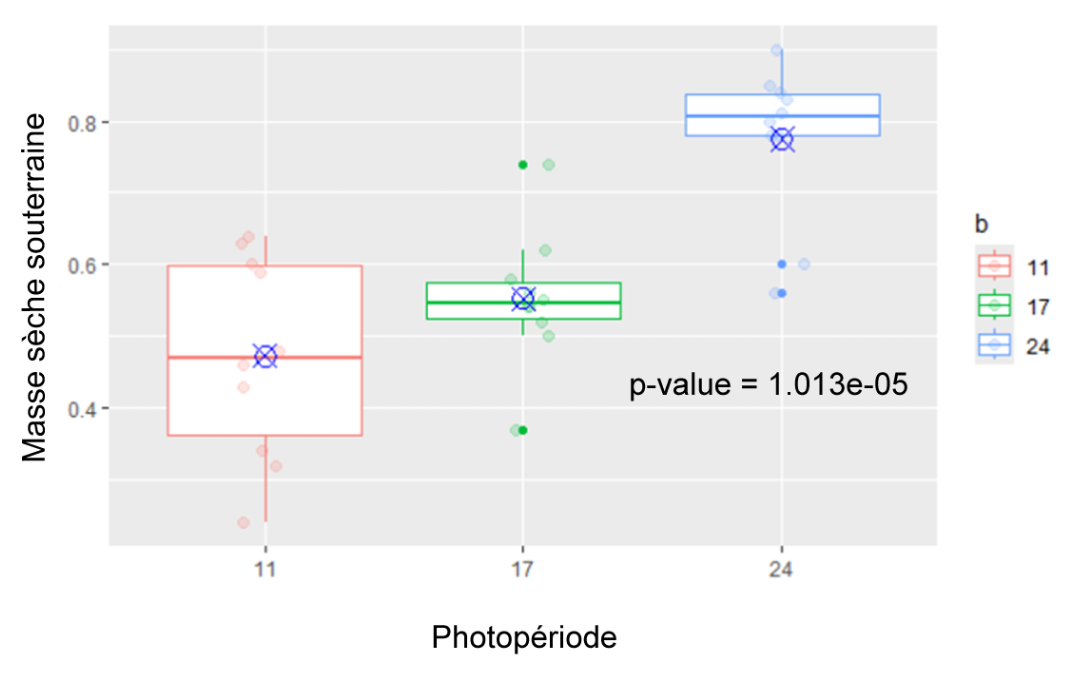
Figure 9 : Masse sèche souterraine des plantes des différents groupes (11h, 17h, 24h) mesurées au 31ème jour
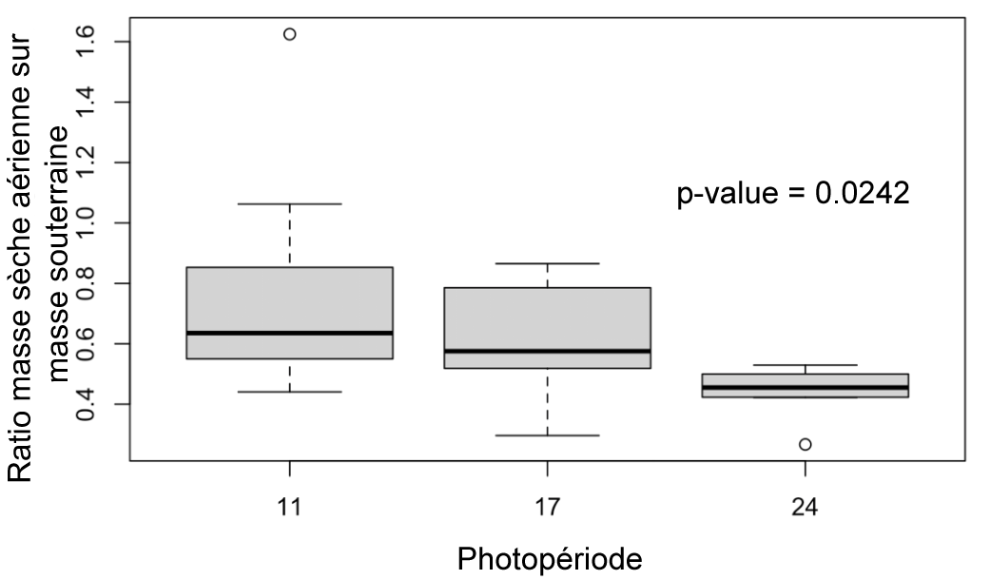
Figure 10 : Ratio des masses sèches aériennes sur souterraines des plantes des différents groupes (11h, 17h, 24h) au 31ème jour
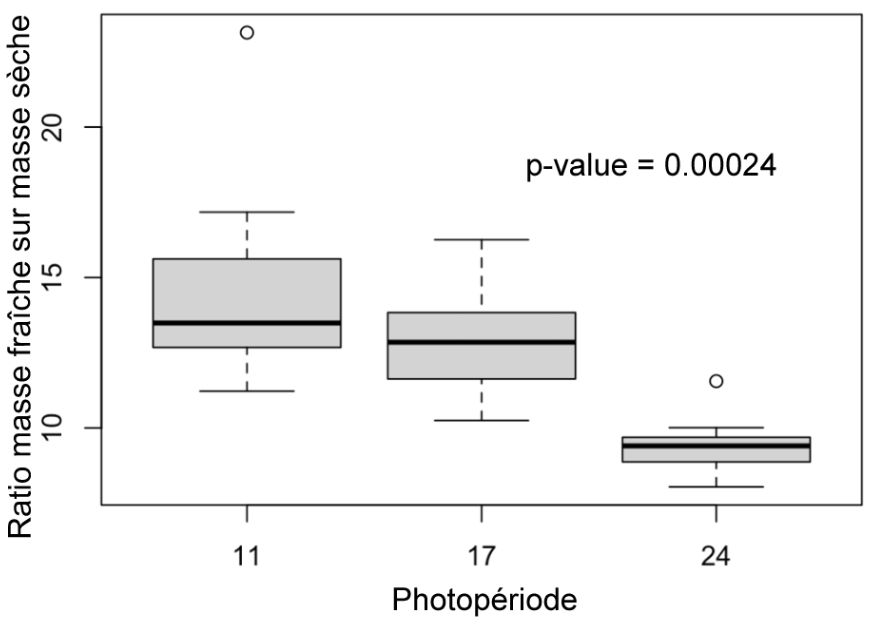
Figure 11 : Ratio des masses fraîches sur les masses sèches des plantes des différents groupes (11h, 17h, 24h) au 31ème jour
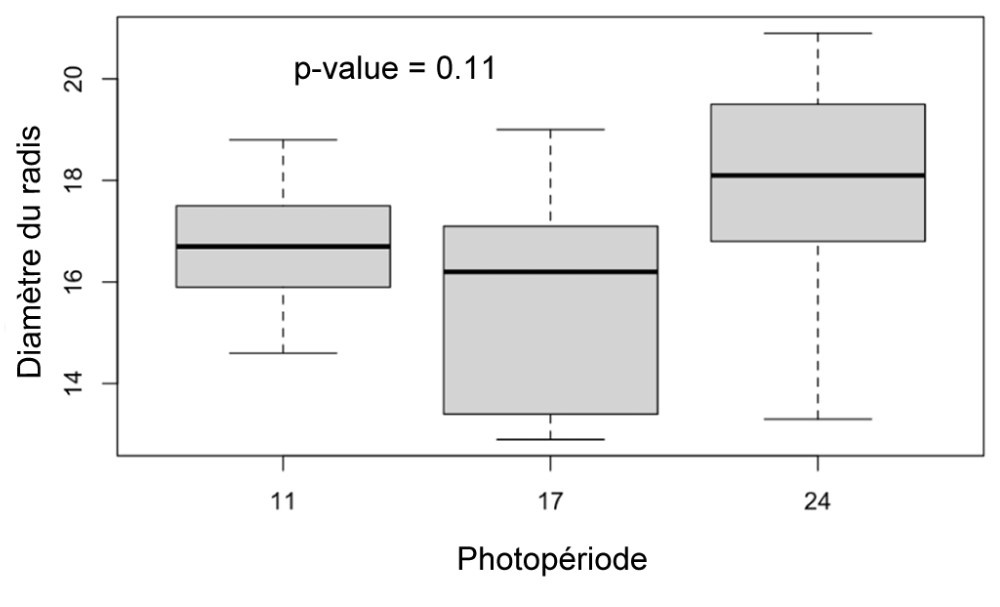
Figure 12 : Diamètre des radis des plantes des différents groupes (11h, 17h, 24h) mesurés au 31ème jour.
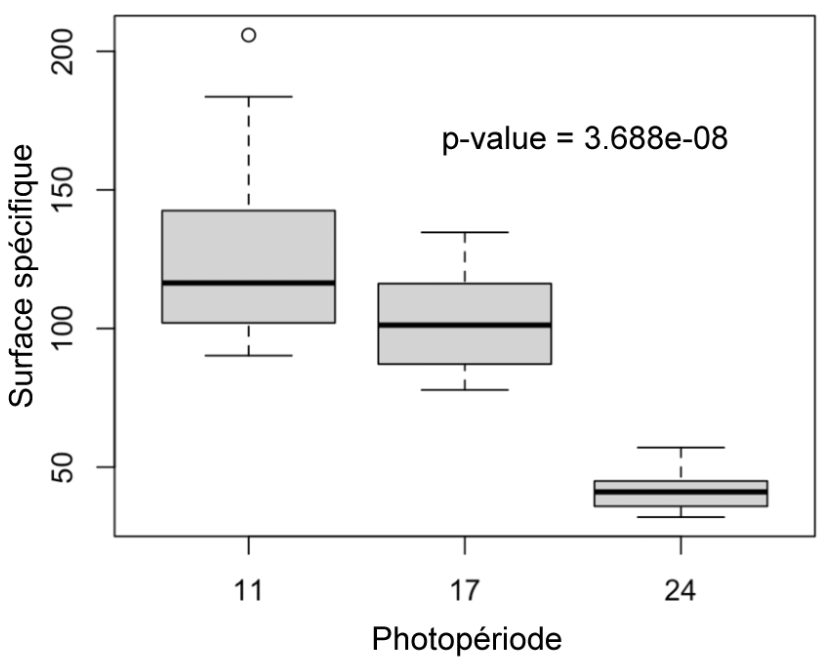
Figure 13 : Surface spécifique des plantes des différents groupes (11h, 17h, 24h) mesurés au 31ème jou
No Comments